All cells, regardless of their specific or specialized functions, perform common activities necessary for life. These vital or housekeeping functions include the production of energy; synthesis and modification of proteins, lipids, and carbohydrates; movement of substances into and out of the cell; changes in or maintenance of cell shape; and duplication the genetic material. In most cases, these activities are performed by specific structural components within the cell. The purpose of this laboratory is to examine these structural components, as detailed by the transmission electron microscope.
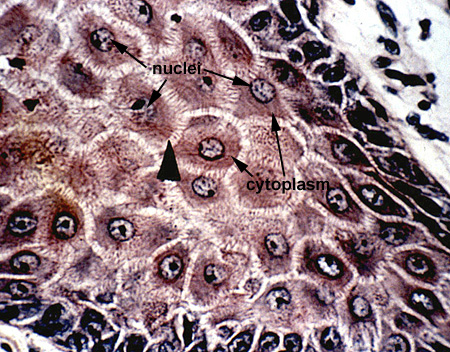
Basically the cell can be divided into two parts, the nucleus and the cytoplasm (Cell). The nucleus is a membrane bound structure containing the genetic material (DNA and associated proteins = chromatin), RNA and forming ribosomal subunits (nucleolus), and the amorphous "nuclear sap" containing soluble material (i.e. enzymes). The cytoplasm is the home of the organelles, which perform energy requiring activities, and inclusions which represent storage materials within the cell. Surrounding these components is the cytoplasmic ground substance or cytosol (Cell 2).
Many of the organelles and inclusions of the cytoplasm are membrane bound structures, that is, they are surrounded by a membrane. These membranes constitute intracellular microcompartments in which enzymes, substrates, or other substances can be segregated or concentrated. Many of these membrane structures are convoluted or folded to provide an increase in the surface within the cell on which physiologically important reactions occur. The membranous organelles of the cell include:
Not all organelles of the cell are membrane limited. The nonmembraneous organelles of the cell include:
The plasma membrane of the cell is not visible with the light microscope. Even so, early cytologists realized the presence of some boundary structure that was essential for the integrity of the cell. It is now known that the plasma membrane is more than a simple retaining boundary of the cell and participates in many functions of the cell. Using the transmission electron microscope (TEM), the plasma membrane has a trilaminar appearance which has been designated as the "unit membrane" (unit membrane). The trilaminar structure is visualized as a dense outer layer, a clear staining middle, and dark staining inner layer. The entire structure measures 8 to 10 nanometers in thickness. The current molecular explanation for the organization of the plasma membrane is the "fluid-mosaic model". This model predicts the membrane consists of a phospholipid bilayer and globular membrane proteins.
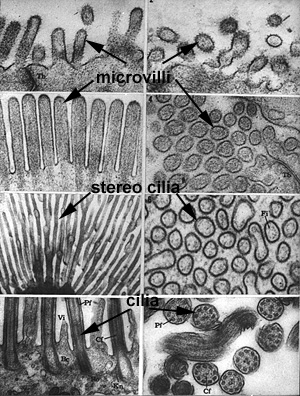
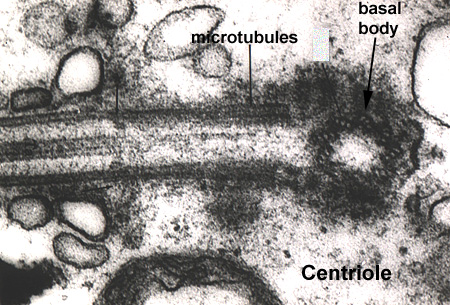
The plasma membrane of the cell can be organized into specialized structures to assist in its function. In cells where the absorption of materials is the primary function (i.e. the intestinal tract), the cell will increase its surface area by membrane folds called microvilli. Microvilli are best described as finger-like folds containing wispy contractile microfilaments (Membrane 1). In cells where the movement of materials is important (i.e. cells of the respiratory tract), the membrane contains specializations called cilia. Cilia share the same finger-like morphology as microvilli, however, they have a much more complex internal structure. Cilia develop from a basal body which is synonymous with a centriole (Centriole - MT2L). Cilia form by the elongation of microtubules within the centriole which form the core or axoneme of the cilium (Membrane 1). Modifications of cilia, stereo cilia, are found in the male reproductive tract and, are often branched and lack microtubular cores (Membrane 1).
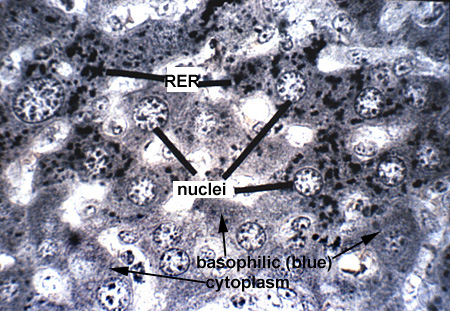
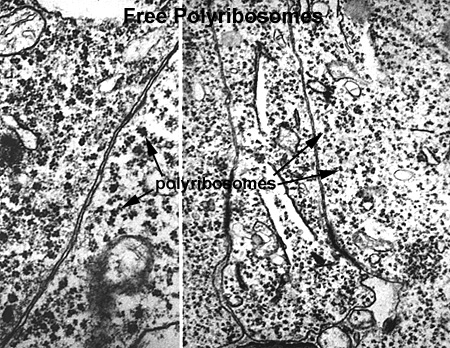
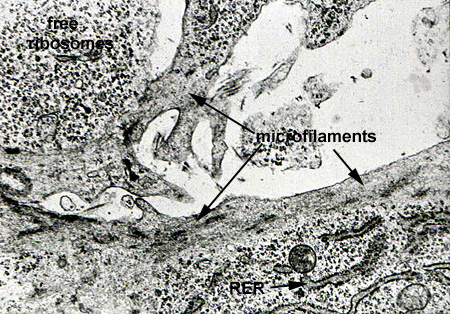
Ribosomes are the cellular organelle responsible for the translation of the mRNA and assembly of the amino acid chain during protein synthesis. Ribosomes can lie free in the cytoplasm or can be associated with membranes (RER) to produce secreted proteins. Free ribosomes are responsible for the synthesis of cytosol proteins and organelle subunits. They can exist as single free ribosomes or in clusters known as polyribosomes or polysomes. By light microscopy they produce a general low level cytoplasmic basophilia, with more localized and intense basophilia associated with RER (Ribosome 1). With the TEM, ribosomes appear as small (25 nanometer) electron dense particles in the cytoplasm. Polysomes can appear as irregular clumps or organized into rosettes or whorls (Ribosome 2 and Ribosome 3).
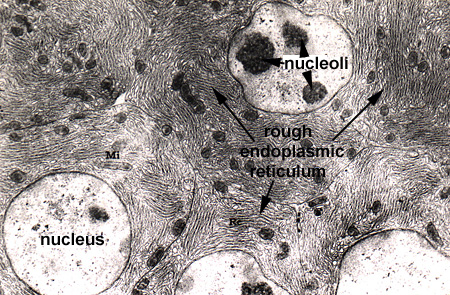
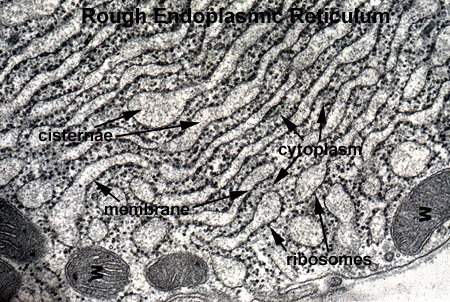
The RER is the cytoplasmic organelle used to synthesize proteins for secretion from the cell. This structure works with the Golgi system to modify and package proteins. In highly secretory cells, it fills the cytoplasm as a lacey network (RER 1). The structure consists of two component parts: a flattened membrane sac which is studded with ribosomes on the outer (cytoplasmic) surface. Newly synthesized proteins accumulate within these sacs in the cisternae which can appear greatly dilated due to the accumulation of product (RER 2 and RER 3).
The proteins in the cisternae of the RER are delivered to the Golgi apparatus (Golgi and Golgi 1). This process is accomplished by small vesicles (transfer vesicles) budding off the RER, moving to and fusing with the Golgi. The unit structure of the Golgi is a flattened membrane vesicle called a saccule. The golgi saccules are saucer shaped, stacked upon one another, and laterally connected. The stack of saccules of the Golgi has two faces: (1) the forming or cis face, which is usually convex and (2) the mature or trans face, which is generally concave (Golgi 2). The forming face is so named because it is produced by the continual fusion of transfer vesicles from the RER to the Golgi. The protein emptied into the Golgi travels through the cisternae where it is modified by enzymes which add carbohydrate, phosphate, etc to the peptide chain. (Note that the Golgi is also the main source of carbohydrates produced by the cell). Once modification is complete, the protein is packed into vesicles which bud off the mature face of the Golgi and travel to the cell surface for secretion (Golgi 3).
Lysosomes are found, in various numbers, in all cell types. They are membrane vesicles approximately 0.5 microns in diameter. They contain several hydrolytic enzymes (proteases, nucleases, glycosidases, lipases, and phosphatases). In healthy cells they serve an important digestive function. Lysosomes are produced by the Golgi and when they bud off the mature face are called primary lysosomes. In routine TEM preparations they appear as homogeneous, electron translucent vesicles (Lysosome 1). When a primary lysosome fuses with a vesicle containing unwanted material it is now called a secondary lysosome. As the source of unwanted material differs, so does the morphology of a secondary lysosome. In general, the homogeneous contents are replaced by the now apparent unwanted material (i.e. bacteria, degenerated organelle, etc). The hydrolytic enzymes of the lysosome then digests the material. Whatever is left over is termed a residual body (Lysosome 1).
SER differs from RER in two ways: first, the outer surface of the membrane is not studded with ribosomes and second, SER consists of membrane tubules arranged in an anastomosing network, not as large, flattened cisternae. SER carries out a number of important functions including the synthesis of lipids, drug detoxification, calcium storage, and glycogen formation.
Oxidation of nutrients and the production of energy occurs within the mitochondria. By TEM, mitochondria are found to be composed of two membranes (outer and inner) (Mitochondria). Both exhibit the typical unit membrane morphology. The outer membrane plays a role in controlling the movement of substances into and out of the mitochondria, uptake of substrates, and release of ATP. The inner membrane is arranged in shelf-like folds called cristae (Mitochondria 1). (Note: Mitochondria are long and often tortuous in shape. They therefore have many different appearances based on the orientation of the section through the structure.) The enzymes associated with electron transport and oxidative phosphorylation are located on the cristae, associated with "toadstool" like structures, the elementary particle. These structures are visible in high magnification micrographs of negatively stained specimens (Mitochondria 2).
Every cell contains specialized cytoplasmic proteins which serve as a support and contractile system, maintaining or changing cell shape. The cytoskeletal structures include the microtubules, microfilaments, and intermediate filaments.
Microtubules
Cytoplasmic microtubules appear as 25 nanometer tubular structures and are readily assembled and disassembled from cytoplasmic pools of the protein tubulin. Microtubules are fairly rigid and play a role in the maintenance of cell shape (Microtubules 1). They are associated with cilia formation (from basal body) (Microtubules 2), and spindle apparatus formation (from the centriole) during cell division (Microtubules 3).
Microfilaments
Microfilaments are fine, thread like structures about 6 to 7 nanometers in diameter. An analogy sometimes employed to indicate function is that the microtubules act as the "bones" of the cell, whereas the microfilaments act as the "muscles" since they provide for movement and shape change. Microfilaments are composed of the contractile protein actin and represent a primitive contractile system, forming large bundles called stress fibers (Microfilaments 1). In most cells, the microfilaments are found in a band just under the plasma membrane (Microfilaments 2).
Intermediate Filaments
Intermediate filaments are generally 8 to 12 nanometers in diameter and biochemically are a heterogeneous group (keratan, dermatan, desmin, and vimentin are a few of the biochemical species of intermediate filaments). The intermediate filaments are not contractile and serve exclusively in a supportive role. They are frequently grouped into delicate bundles (fibrils) in the cytoplasm, suited to meet stress and provide an overall girder-like support system.
Inclusions are "nonliving" components of the cells; they do not participate in the energy-requiring reactions. They represent stored materials within the cell. Of the three basic foodstuffs, only carbohydrates and fat are stored by cells. Neither is found enclosed by a membrane. Carbohydrate is stored as glycogen deposits in the cytoplasm. It appears as electron dense granules, slightly larger than ribosomes, and is often organized into rosettes. Fat is stored as droplets within the cytoplasm. The other major type of cell inclusion is the membrane bound pigment granule. Melanin is a brown-black pigment found in the skin and eyes.
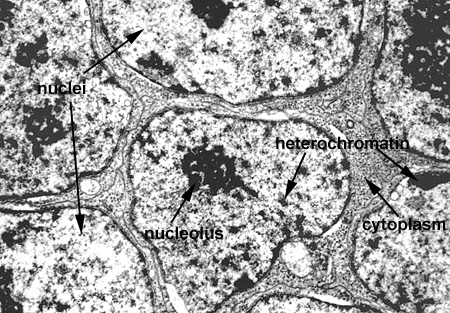
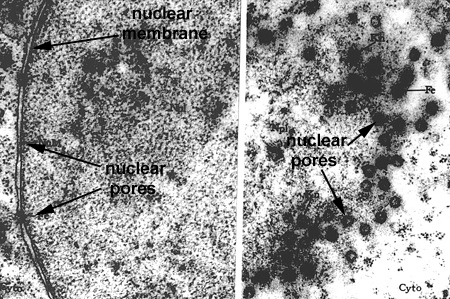
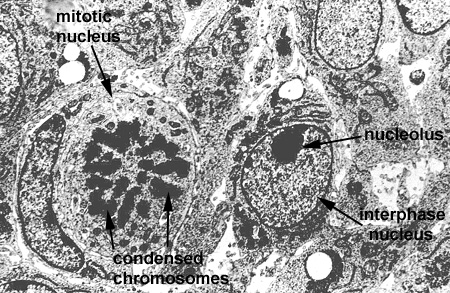
In this laboratory we will study the structures of the "interphase nucleus", a nucleus not in the process of mitosis. With the light microscope, the nuclear appears limited by a dark blue staining line, the nuclear envelope or membrane. Within each nucleus there is one (or more) large blue staining bodies, the nucleolus (or nucleoli). There are also numerous, poorly defined aggregates of blue staining material which is referred to as chromatin. The rest of the nucleus (pale staining) is called the nuclear sap (Nucleus 1). By TEM, the nuclear envelope consists of two membranes (each 8 nanometers thick) and the heterochromatin and nucleoli as electron dense granular material (Nucleus and Nucleus 2). The outer membrane is connected to the RER of the cytoplasm. The envelope contains numerous, octagonal nuclear pores which allow for nuclear:cytoplasmic interaction (Nucleus 3 and Nuclear Pores). Chromatin is a complex of DNA and its associated proteins. It is found in various degrees of condensation. The most condensed form is formed is found during cell division as the individual, supercondensed chromosomes become visible. In the interphase nucleus, the chromatin is found in an uncondensed and active form (euchromatin) or a condensed and inactive form (heterochromatin) (Nucleus 4).
The nucleolus is the site of ribosomal RNA (rRNA) and ribosomal subunit production. It is composed of extrachromosomal rDNA, rRNA, and ribonuclear proteins. By TEM it consists of light and dark areas. The dark areas consist of the pars granulosa (granules of rRNA and proteins) and the pars fibrosa (filaments of rRNA). The pars fibrosa represents the darkest regions and is found surrounding a core of less dense material, the fibrillar center. The fibrillar center represents the extrachromosomal rDNA (Nucleolus).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}